|
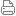
生物氧化(三) 7. 在氧化呼吸鏈中的NADH-Q還原酶、細胞色素還原酶和細胞色素氧化酶催化的三步反應中,自由能的變化都足以將H+從線粒體內膜基質“泵”出到線粒體的內外膜間隙,產生氫離子梯度,為下一步產生ATP準備所需的自由能。 (3) 電子傳遞的抑制劑 為能夠阻斷呼吸鏈中某部位電子傳遞的物質,可用來研究電子傳遞順序。 常見的抑制劑有: 1. 魚藤酮,安密妥 阻斷NADH-Q還原酶內的電子由NADH向CoQ的傳遞。 2. 抗霉素A:抑制細胞色素還原酶中電子從QH2到Cyt c1的傳遞。 3. 氰化物,疊氮化物與a3中血紅素Fe3+作用。CO與a3中Fe2+作用,均阻斷電子在細胞色素氧化酶中傳遞作用。上述各種抑制劑的抑制部位可表示為NADH → NADH-Q還原酶 -║→ QH2 -║→ CytC1 → CytC → 細胞色素氧化酶 -║→ O2 魚藤酮, 抗霉素A CN-,N3-,CO 安密妥
3、氧化磷酸化作用 (三)氧化磷酸化作用 將生物氧化過程中釋放的自由能用以使adp和無機磷酸生成高能ATP的作用都在細胞線粒體內膜發生作用。 氧化磷酸化全過程方程式為: NADH+H++3ADP+3P1+1/2O2→NAD++4H2O+3ATP (1) P/O比:一對電子通過呼吸鏈傳至氧所產生的ATP分子數。 NADH的P/O比為2.5(過去為3),從呼吸鏈NADH-Q還原酶進入。 FADH2的P/O比為1.5(過去為2),是從細胞色素還原酶處進入電子傳遞鏈。
(2) ATP合成部位 為三個能量釋放部位。 一對電子經NADH-Q還原酶,細胞色素還原酶和細胞色素氧化泵泵出質子數分別為4,2和4。合成一個ATP要3個質子通過ATP酶驅動合成2.5個ATP需7.5個質子驅動,合成1.5個ATP需4.5個質子驅動,多余的質子可能用于將ATP從基質運往膜外細液。 ATP合成是在線粒體ATP酶作用下完成的,現稱ATP合酶,又稱復合體V。
(3)能量偶聯假說 電子傳遞釋放出的自由能和ATP合成是與一種跨線粒體內膜的質子梯度相偶聯的,1961年提出,1978年獲諾貝爾化學獎。 電子傳遞鏈為一個H+泵,使H+從線粒體基質排到內膜外;內膜外H+比內膜高,形成H+濃度梯度;電化學電勢驅動H+通過ATP合酶F0F1ATP回到線粒體基質,釋放自由能與ATP合成偶聯 此過程為: 1.質子泵出需要能量:由電子流過復合體Ⅰ、Ⅲ和Ⅳ獲得。 2.質子泵出使細胞溶膠側H+濃度提高,產生膜電勢。 3.膜電勢驅動質子流通過ATP合酶,同時釋放出與酶牢固結合的ATP,ATP是在釋放自由能驅動力下ADP與Pi合成的。 F0F1-ATP酶:F0單為嵌入線粒體內膜中,為質子通道;F1在膜外為球狀體。催化ATP的合成。
(四)氧化磷酸化的解偶聯和抑制 一般情況下,電子傳遞和磷酸化是緊密結合的,但特殊的試劑可將氧化磷酸化過程分解成單個的反應。 (1)解偶聯劑: 只抑制ATP的形成過程,不抑制電子傳遞過程,使電子傳遞產生的自由能變成熱能。如DNP(2,4-二硝基苯酚),當pH=7時,DNP呈酚負離子形式,不能透膜。在酸性環境(H+濃度提高),DNP接受質子而成酚分子形式,脂溶性而易透膜,將H+帶入膜內,破壞了跨膜梯度的形成,故DNP又稱質子載體。 褐色脂肪細胞線粒體內膜上有特殊H+通道,H+流回不經過F0F1-ATP酶,不產生ATP產生熱,維持體溫。
(2)氧化磷酸化抑制劑: 因抑制ATP形成而使電子傳遞停止,如寡霉素。但為寡霉素抑制的電子傳遞會由于加入DNP,使H+濃度差消除,而使電子傳遞恢復。
(3)離子載體抑制劑 它們與K+等離子結合,作為離子載體穿過膜,消除膜電勢,如短桿菌肽,纈氨霉素ATP不能生成。
| 



 發表于 2022-8-1 16:49
發表于 2022-8-1 16:49
 收藏
收藏 分享
分享 支持
支持 反對
反對